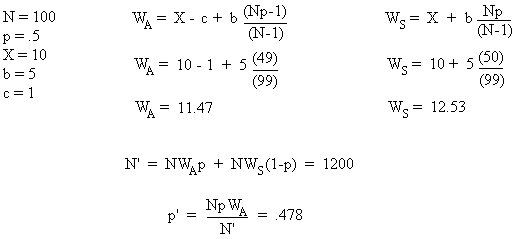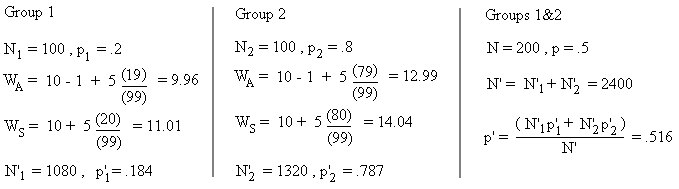本条目初稿作者:陈安慧
本条目短网址: http://appedia.arc.capn-online.info/altru
对于人类的本性,历史上很多学者有自己的观点,中国传统思想主要代表的道家和儒家学者均认为人性本善[7][8]。西方科学界也有对这个问题的探讨。在革命性理论中,达尔文的《演化论》是不可忽视的一个。他提出了生物界的自然选择学说,描述自然界对一个种群世代的基因构成造成影响的过程。首先,个体内部的基因变异是由DNA复制过程中的错误引起的。这些变异中的部分会使得个体特征发生极大改变。那些可以提高后代成功率的特征肯定会越来越多地在后代中出现,从而逐渐消除那些对个体的生育能力不利的特征。随着世代相传,这个过程又会大大改变种群内部拥有这种特征个体的数量比,致使这一种群更能适应特定的环境。
尽管达尔文的理论成功地解释了各种各样的自然现象,但把这一理论应用于动物利他主义的演化时却在强调人个体利益的西方世界导致了较大争论[1]。在争议中基因选择理论脱颖而出,成功的说明利他主义的诞生是符合演化论的。利它行为是一种长期演化积累下来的生物本性。本文讨论各种相关理论,除了基因选择理论外,其他理论由于简单化看待问题,往往无法成功的解释大量现存的生物利他行为。
基因选择理论同时也为人类和动物间的跨物种利他行为提供了良好的解释:人类和其他动物共享大量的共同基因。
争论
利他主义,指个体有意牺牲自己的部分生殖能力来提高另一个体的生殖能力。这种生殖能力经常被理解为个体的基因适合度;更精确的定义为,相对于种群中其它个体来说,此个体对后代基因库的贡献。因此,利他主义者就是通过降低自己基因适合度来提高他人基因适合度的个体。一些例子可以帮助
我们更好地理解利他主义:非洲猎狗群体出动去捕食时,一只猎狗自愿留下当“保姆”照顾群中的小狗;当鹰靠近时,一只鸟发出警报声,导致鹰将注意力集中到自己身上;某人跳进游泳池去救一位溺水的陌生人等。从利他主义者的立场出发,很明显,这些行为需付出的牺牲程度不同,但从总
体上来说,以上所有个体都会减少预期中的后代数量。[1]
由此,有些人认为利他行为存在一个悖论。从定义上看,如果利他主义会降低自身适合度,利他特征就是达尔文自然选择学说中所谓的不利特征,而且最终会在一个种群中被彻底消除。即使一个种群在存在初期拥有一些利他主义者,也很容易从内部将其根除,因为那些自私的个体会利用利他主义者帮助他人的倾向,从而使利他特征最终灭绝。[1]
西方世界早期的演化论讨论忽略了利他主义,但最近几十年里,在争论自然选择究竟在哪一个层次上运行——是在基因、个体、亲属群,还是整个种群层次上时,利他主义成了这一争论的中心议题。大量的理论被提出来解释这一问题,下面是其中的几个。[1]
群体选择
使利他主义可以演化的第一个机制是群体选择,以下威尔逊(E.O. Wilson)的模式是一个例子。假设一个种群中有N个个体,分为两种类型:利他类型(A)和自私类型(S),两种类型各占总数的比率为p和(1-p)。由于A类个体的利他天性,群体中成员的后代数量将会增加,这个增加量可以用b(相对的后代增加量)以及群体中可用利他个体的百分比的乘积来表示。对S型来说,这个值是b×[Np / (N-1)],而A型是b×[Np-1 / (N-1)]。
这一差别表明利他主义者并不能从自己的利他行为中受益。如果我们再设值c(利他者由于自己行为而减少的后代数量)和x(每个个体在不行使利他行为的前提下预期的后代数量),再把存在利他行为前提下,两类个体预期的平均后代数量设为w。
公式图片来源[1]
由于b和c总是正数,N总是大于1,很快可以看出,在一个固定的群体中,自私个体的后代总是多于利他的个体。下面给出一个有具体数字的例子,假设群体成员总数为100,其中一半为利他者,一半为自私者,N′是下一代个体的总数量,p′是下一代中A型个体的总数量。
公式图片来源[1]
从此例可看出,利他个体的比例从第一代的50﹪降到了第二代的48﹪。用此计算方式一代一代算下去,我们会发现利他个体在群体中所占比例很快会变得非常小。如果这一种群的个体总数已接近于其环境容量(所处自然环境对某生物的最大容量),且总数量稳定,那么从此例可推断,此种群内部的利他特征最终将完全消失。这一数学公式又使我们想到了前面提过的悖论——如果利他主义处于自然选择的不利面,它又怎么能世代遗传下来且得到演化呢?[1]
将这一数学公式做一个小小的调整就能解决这一悖论。这次,我们设两个个体数目相同的群体,但它们内部的利他者和自私者所占的比例不同,计算方式如下,其中X、b、c的值与上例相同:
公式图片来源[1]
虽然分开来看两组中的利他者所占比例都在减少,但如果把两组合在一起,利他者的总体比例却在上升。产生这种反直觉的结果是因为,一个群体中拥有的利他者越多,它的繁殖能力就越强。因此,如果一个物种拥有多个群体,且群体间的利他者与自私者数量比例各不相同,只要这些群体偶尔因生育的需要聚在一起,利他主义者的数量就会无限增加。尽管这一公式说明了利他特征是如何能够增加的,但它不能解释利他主义最初是如何产生的。为此,我们必须把注意力转向另一类型的选择。[1]
亲缘选择
一些人认为达尔文演化论的困境之一是无法解释社会性昆虫中只负责工作的不育阶层的延续。如果这些不育阶层无法留下后代来继承它们的特征,它们如何进化呢?自然选择学说中的一个特殊概念——亲缘选择消除了这一困境。亲缘选择的最基本假设是:个体要想最大化的在后代中传播自己的基因,既可以通过提高自己的基因适合度,也可以通过提高其亲属的基因适合度,因为亲属与其自身有着大量的相同基因。要充分理解亲缘选择,首先要了解亲缘系数和广义适合度。简单说,两个个体间亲缘系数就是他们从共同祖先那里继承下来的共同基因的概率。比如说父亲或母亲与子女的亲缘系数是1/2 ,因为子女会从父亲或母亲身上各继承一半的基因。亲兄弟姐妹之间的亲缘系数也是1/2 ,祖父母与孙子女间的是1/4 ,第一代堂兄弟姐妹或表兄弟姐妹间的是1/8。广义适合度是指,个体自身适合度的总量加上其所有亲属相关适合度的总量(威尔逊,1980)。因此,虽然不育昆虫的自身适合度为零,如果它用自己的生命来帮助亲属,它的广义适合度就会相当高了。[1]
现在我们可以用亲缘关系和广义适合度的概念来判断自然界中何时会出现亲属间的利他主义现象。比如假设你是一只画眉鸟,突然注意到有一只老鹰正在飞近,如果你向周围的同类发出警告声,即使警告声向老鹰暴露了你所在的位置,并最终导致你的丧生,但你的警告声可以拯救周围适当数量的近亲,这样,在自然选择的压力下,你的那个(或那些)使你发出警报叫声的基因仍能成功地被继承下去,因为你的广义适合度比那些不向亲属们警告危险的个体要高。但是,究竟怎样理解适当数量的近亲呢?[1]
英国生物学家汉密尔顿(W.D. Hamilton)以数学方式解决了这一问题,得出了汉密尔顿法则。根据汉密尔顿法则,为了使两个体之间有利他行为的出现,必须满足k﹥(1/r)的条件,k指亲属获得的适合度与利他主义者失去的适合度的比率,r是亲缘关系程度。也就是说,一个个体会愿意献出自己的生命来救助自己的两个兄弟(因为r=1/2),或救助四个孙子女(r=1/4),或救助八个堂兄弟姐妹(r=1/8)。以上列举的事例很极端,因为利他主义者(由于死亡)个体适合度降到为零。除此外,汉密尔顿法则还可以解释利他主义中一些更微妙的情况。[1]
为了更好地理解,我们来看下面一个例子。假设你是一只在森林中寻找食物的兔子,你发现了九根胡萝卜,但只能吃得下四根,每吃掉一根胡萝卜就可以得到任意两单位的收益。如果你守住秘密,独自吃掉胡萝卜,你的总收益就会是4×2=8个单位。但是如果你的兄弟和你兄弟的朋友(与你无亲缘关系)就在附近,你把他们喊来与你分享,胡萝卜在你们三者之中平分,那么,总体上说,你基因的广义收益将是(3×2)(1)+(3×2)(1/2)+(3×2)(0)=9个单位。除了你表面损失了一根胡萝卜和非亲属得到了三根胡萝卜外,在这一事件中,你的基因仍因与他人分享食物而受益。[1]
图1.图片来自[1]
有人或许会问,一只兔子怎么能够在邻近的众多兔子中识别复杂的亲属关系呢?它怎么能够在具体的情形下进行必要的数学运算来决定是做出还是不做出利他行为呢?答案很明显,它不能。基因没有遥感能力,不能在其它个体上探寻与自己相同的基因。然而生物体能够对那些似乎拥有相同基因的个体更友善。有多种方式可以使他们做到这一点。首先是理查·道金斯(Richard Dawkins)提出的“绿胡须效应”。如果一个基因使其携带者生有显著的生理特征(比如绿胡须),而且这个携带者又能对其他拥有相同生理特征的个体表现友善,那么长着绿胡须的个体之间的利他主义就可以演化。由此可以看出个体没有必要区分他们的相同基因是源于共同的血统,还是仅仅为无规律变异的一个巧合。因为从利他基因的角度来看,做这种区分只是浪费时间。虽然绿胡须效应在理论上可行,但道金斯本人也承认,单个的基因未必能既生出迥异的生理特征(如绿胡须)又具有利他主义倾向。[1]
个体更愿意对有相似基因的他人做出利他行为的另一表现是,与亲属建立亲密关系。母亲们当然都很清楚自己与子女的关系。亲兄弟姐妹之间也很明白彼此间是有亲缘关系的,因为他们是在一起成长起来的。由此而产生的识别亲属的办法是通过个体间居住地的远近。比如你是某个群体中的一员,而且很少走出居住范围,那么很有可能你周围的邻居都是你的亲戚。这样一来,利他基因不一定只针对亲戚,而更可能不加区别地针对任何一位近邻。这一机制可以用来解释猴群和鲸群内部的利他主义行为,因为这些群体中的任意两个个体之间都可能有很亲的血缘关系。[1]
虽然这些类似的亲属识别方式是亲缘选择的主要基础,但不可忽视的事实是,在并不存在环境因素(如两个体比其它个体相隔更近等)的影响时,有时近亲对非亲属更友好,人们至今还无法解释这种现象。[1]
互惠利他主义
虽然亲缘选择为自然界的大多数利他行为提供了有力的解释,但无法解释两个无关系的个体(尤其是高级哺乳动物)之间的利他行为。为此,特里弗斯(Robert Trivers)提出了互惠利他理论,指个体为帮助另一方做出利他行为是为了日后对方会有所回报。这种交换的结果是,双方都提高了自身的适合度。[1]
假设一人落水,附近一名与他无任何关系的女性赶来救他。如果得不到帮助,这个人溺亡的可能性是1/2,如果那位女性决定救他,她自己溺亡的可能性就是1/20。有人可能倾向于下结论说这位妇女不会去实施救助行为,因为这样做的直接后果是既降低了她的个人适合度,也降低了她的广义适合度(因为这个男人并不是她的亲戚)。然而如果日后某一天,这位妇女落水,那位男性又正好在附近,他很可能会去救她,以回报当初妇女救自己时所冒的同样的风险(1/20)。现在我们可以看出,两个人都在用1/20的死亡几率换1/2的死亡几率。因此,有着众多参与互惠交换的利他主义者的种群,能最终提高他们总体的基因适合度,而那些摈弃利他主义、充满竞争的种群却不能。[1]
但这个理论中也存在一个问题。上例中的那个男人,在已经获救后为什么还要去实施互惠式的回报,为什么他在日后还要冒那些不必要的风险?在人类社会中答案很简单:欺骗者会受到惩罚,不仅来自于他没有提供回报的那个人,更来自于广大的社会。欺骗者由于欺骗行为导致的适合度降低,比他实施回报导致的适合度的降低要大得多,而且他还会处于自然选择机制中的不利面。然而大多数的物种不可能具有这种识别和记忆能力,这就是自然界中互惠利他比基于亲缘选择的利他要相对较少的主要原因。[1]
互惠利他概念与博弈论中著名的“囚徒困境”所用的针锋相对策略有紧密的联系。在博弈中,选手两人一组,可选择下列两种策略的其中之一:
(1)合作,(2)背叛。各博弈选手所获利益(在此文中这种利益可设为自身生殖适合度的增加)不仅取决于自己所选的策略,同时也取决于对手的策略。所获利益值如下矩阵图所示(图中的具体数字并不重要,重要的是数字的不等)。
| | | 选手A | 选手A |
| | | 合作 | 背叛 |
| 选手B | 合作 | 11 | 0 |
| 选手B | 背叛 | 20 | 5 |
选手A在生殖适合度上所获利益值只要两者一旦被置于互斗地位,很明显最佳选择应是背叛——无论对手怎样做,背叛都比合作的结果好(如果对手合作,自己背叛与合作的获利比为20:11,如果对手背叛,这个比值为5:0)。因此如果选手们只博弈一次,他们不会选择合作——自然选择更倾向于背叛者,任何一个合作者最终都会在群体中被淘汰。然而如果选手们反复多次地互斗,而且会根据对方以前的选择来调整策略,情况就会复杂多了。在这种所谓“重复的囚徒困境”中,每次都背叛并不一定是最好的选择。事实上,阿克塞尔罗德(Axelrod)和汉密尔顿(Hamilton) (1981)指出只要将来两者再相遇的概率很高,针锋相对策略就会产生很高的利益。采用针锋相对策略的选手要遵循两个基本原则:(1)在第一个回合选择合作策略;
(2)此后则选择与其对手在前一个回合所选择的策略相同的策略。如果一个群体中的所有个体都采用针锋相对策略,那么就不会有诸如“每次都背叛”这样的可供选择的策略了。因此针锋相对策略是一种“演化稳定策略”(Maynard Smith (1982))。
显然这个结果对于利他主义的演化是很有意义的。囚徒困境中的合作就相当于利他性行为,背叛相当于自私行为。我们直觉地认为,当有机体期待将来获得回报时会认为选择利他行为更有利,阿克塞尔罗德和汉密尔顿的研究成果为这一直觉想法提供了有力的依据。只要有机体之间存在着各种各样的联系,而且能根据其它有机体的以往行为来调整自己的行为,理论上,互惠利他主义就可以演化。[2]
基因选择
基因选择的观点因Richard Dawk的《自私的基因》一书名声大噪,基因选择论被认为是最佳的自然选择理论,因为它解决了其他选择理论(群体选择、亲缘选择等子理论)不能解释或者有困难解释的自然现象,利他主义就是其中一列。简单的说,该理论认为,‘自私’的单位不是生物的个体,而是基因。生物个体是基因的载体,其任务是从长期的角度最大程度扩大所携带基因在自然界里存在的数量。一个例子,人们之所以最多的帮助直系亲属是因为直系亲属和我们分享最多的基因。所以人的利他行为变得很容易解释的——他人乃至很多动物都和人类共享很多基因。所以可以说利他主义是人类的本性,且是符合演化论的原理的[4]
录像1:Richard Dawkins介绍《自私的基因》出版30年
http://www.youtube.com/watch?v=ciFe8JmR-nY
报警
图2. 图片来自[4]
也许在动物的利他行为中,讨论得最多的例证是它们的报警行为。当捕食者靠近,而这一区域生活的被猎食动物不止一只时,第一个发现捕食者的个体会发出警报声以提醒它的同类们,同时将捕食者的注意力吸引到自己身上来。很明显,在这种情况下引起捕食者的注意对自己是极不利的。
P.W. Sherman积极解释了这一现象,他假定任何警报声都具有以下六个功能中的一个(或几个)。
1. 报警者发出警报声后可能会引起同类的集合或紧张行为,因此,实际上会使得捕食者把注意力从自己身上移开。
2. 被捕食者盯上的猎物可能在听到警报声后意识到自己已暴露,从而吓得失去胆量逃跑。
3. 如果捕食者更乐意去猎取它曾经成功捕获过的物种,那么警报声有可能减少它日后对自己种群的攻击。
4. 如果受益者们做出互惠利他式的回报,个体也许会发出警报。
5. 如果被捕食种群是由多个群体组成,作为群体选择的结果,各群体中具有报警倾向的个体所占比率不尽相同,报警行为有可能出现(类似于前面提到过的例证)。
6. 虽然报警者冒着付出生命的危险,但如果周围的同类中有自己的亲属,报警声就可以帮助亲属们逃跑,从而提高自己的广义适合度。[1]
我们能很快得出结论:第1,2和3条功能不能被划分到利他主义中来,因为报警者同样从自己的行为中获益。第4项功能听起来很合理,但特里弗斯提出了几条反对意见。首先,很难说一群个体如何识别出并歧视不具有互惠利他行为的“骗子”;其次,没有证据证明野生动物会由于附近动物们不付出互惠行为而不发出警报声。第5项功能与前面谈到过的群体选择的数学公式几乎是一致的。这一点可以帮我们更深入地研究第6项功能。[1]
Sherman已证实了松鼠的亲缘选择在利他主义演化中的有力推动力量。在对大约1100只松鼠进行了长达3000小时的观察后,Sherman发现,正处于繁殖期的雌性松鼠更倾向于会发出警报而不是被警告,同时也比那些非繁殖期的雌性松鼠更乐意发出警报,虽然繁殖期和非繁殖期的雄性松鼠间没有差别。Sherman认为这一结果源自于松鼠的母系亲属结构,以及在这一物种中雌性更具有家长作风。[1]
食物分享
图3. 图片来自[1]
威尔逊(E.O. Wilson)指出,除了自杀外,没有比分出自己食物更利他的行为了。很多物种都存在这种形式的利他行为。长臂猿和黑猩猩都会分食物给向其乞讨的同类,非洲猎狗也会在成功出猎后,带回新鲜肉食分给留下照顾幼崽的同伙。在食物分享方面最好的文献研究来自于威尔金森(Gerald Wilkinson)对吸血蝠的研究。吸血蝠以吸食其他动物的血为生,虽然并非每单只吸血蝠每夜都能找到猎物,但一旦它找到了,它就会吸很多血,吸血量会超过自身的需要量。吸血蝠们回到群居地后,成功吸到血的吸血蝠就会将血反哺给那些不太幸运的吸血蝠们。这种分享食物行为经常发生在亲戚之间(通常是母亲与子女之间),但也有很多时候,分享食物的两者之间没有任何的亲缘关系。威尔金森假定,在满足下列三个条件的情况下,这种慷慨行为就应该是互惠利他主义的结果。
1. 吸血蝠群中成员间的交往时间应足够长,以确保彼此间有多次这种交换。
2. 接受帮助的收获应大于捐助付出的代价。
3. 做出过利他性捐助的吸血蝠必须能识别出曾经拒绝付出回报的吸血蝠,并从此不再向它们提供血液。
通过对野生蝙蝠群的简单观察就可以发现,前两个条件是可以满足的。威尔金森证实,第3个条件也同样可以满足。他从一个吸血蝠群中抓出几只,使它们挨饿,其它吸血蝠们回来后都很乐意反哺这几只吸血蝠(这几只吸血蝠或许以前曾与它们分享过食物),但它们就不愿意反哺另一组新加入的吸血蝠,因为它们之间以前没有交往。因此我们可以得出结论,吸血蝠间的食物分享行为至少有一部分是互惠利他的结果。[1]
人类的利他主义
互惠利他的基因演化模式很难解释在自私的同类群中利他基因最初是如何出现的。虽然学者们提出了一些可能的机制,但有这样一个观点:大多数的利他行为都始于自私的目的。分享和帮助属于两种不同的行为范畴,不应被混淆。动物的资源分享符合的是自私模式而非互惠利他模式。大量的观察发现,人类互换礼物也是出于自私而非利他。这些都说明,我们需要谨慎看待利他主义的演化模式。[3]
现在有人愿意支持这些利他主义的理论,并把它们运用于人类,尤其认为人类是所有动物中最具利他性的,这种做法是很合乎情理的。利他主义以各种形式存在于社会生活的方方面面,如分享食物、照顾病人、送礼物、甚至通过教育来分享知识。在现代人类社会中,金钱的使用也被看做是互惠利他的一个基本例证,因为金钱本身没有价值,它的价值体现在将来某天某人会为了它而卖给你有价值的物品。
很容易看出,通过亲缘选择,利他主义在人类身上应该已有了怎样的演化。史前的狩猎采集部落几乎都由近亲组成。早期人类因为拥有语言及发展了的智力,比起其它灵长目动物来,更能识别亲属和辨别亲缘系数的微妙不同。J. Rushton证实,人类不仅倾向于对自己的亲属表示亲近,也倾向于对那些有着相同基因特征的人表示亲近,并把这作为通过亲缘选择做出利他行为的证据。通过一系列大范围的研究,J. Rushton证明了,总体上,朋友间的基因相似度大于陌生人之间的基因相似度。他还有另一个重大发现,男女之间互相吸引并非偶然,更有可能是因为彼此有着相似的血液标志物。两个人之间的对彼此的行为表现可能由他们的基因相似度决定,这一理论具有深远意义,而且它所包涵的领域已远远超出了利他主义讨论的范畴。[1]
毫无疑问,在人类社会利他主义的形成中,亲缘选择扮演着重要的角色,但从总体上来说,互惠利他的作用更为显著。人类很容易适应那些使得互惠性利他得以产生的环境:长期维持的关系、积累下来的用以区分互惠者和非互惠者的记忆、惩罚非互惠者的措施等。有一个猜测是,某些人类的复杂情感已经得以演化,来提高互惠利他的机制。感激和同情可以增加一个人从利他主义中得到好处的机会,因为有这两种感情的人向他人施惠的概率很高。同时,内疚感会折磨那些非互惠者,使他们最终决定将来不再拒绝回报。特里弗斯甚至提出了共同演化的复杂模式,用不易察觉的方式来欺骗邻里和提高自己识别隐蔽骗子的能力,这两种压力联合起来,为人类智力的开发和达到目前的高智商做出了贡献。
功利主义的利他
西方世界文化重视个人利益,所以学者在解释利他行为动机的时候,总会从自私为出发点。为迎合文化氛围,道金斯(Richard Dawkins 1976)使用了“自私基因”这一术语来解释利他行为,给公众内造成了对利他行为出自功利动机的错觉。事实是,基因试图使自己在基因库中出现的概率增大当然只是一个隐喻,基因不会尝试去做任何事。演化是物理过程的描述,增大基因总数的生物行为,都将在演化中占有优势[2]
亲缘选择和互惠利他理论能运用于人类行为吗?生物学家赞同智人是演化了的物种,那么通常的演化理论应该都能运用于智人身上。然而人类的行为也受文化的影响,而且通常是有意识的信仰和渴望的结果(虽然这并不意味着基因就对此没有影响)。人类行为的确是符合演化论的。通常人类更倾向于向近亲表现出利他行为,比如帮亲属抚养后代,这正和亲缘选择中所讲的相同。同样,人类也的确倾向于帮助过去曾帮助过自己的人,这一点又正如互惠利他所言。表面上看有大量的人类行为有违演化论的观点。以收养孩子为例,显然不养育自己的孩子而去收养其他小孩会降低做父母的生物适合度,所以收养孩子是一种利他行为,但这种行为并不会使亲属从中获益——因为养父母通常与被收养的孩子没有血缘关系——也不会使养父母从互惠的角度获益多少。所以尽管亲缘选择和互惠利可以帮我们理解某些人类行为,但这两个理论不能运用于所有人类行为中。只有基因选择可以解释这里的悖论。人和陌生人共享大量共同的基因[2]。基因选择同时也可解释人类和动物间的利他行为,人类和其他动物共享大量的共同基因。
索伯(Sober, 1994)认为,即使我们用演化论来理解人类行为,仍然没有明确的理由说明人类不是心理利他主义者而非功利利它者。相反,自然选择可能更偏爱那些真正关心帮助他人的人。假设好好照顾孩子会取得演化上的优势——一个貌似令人信服的观点,那么那些真正关心孩子的父母们,也就是非功利主义利他主义者,就会比那些不想关心或根本不关心孩子的家长们拥有更高的全面的适合度,也就能更多地传播他们的基因。因此演化过程很好地引导了非功利主义的利他的演化。与西方社会常见的想法相反,用演化理论分析人类行为会发现,人类并非只愿被自身利益驱动去做事。可以使基因提高将来的出现概率的一个策略就是,真正的非功利主义的利他。[2]
小结
利他主义是动物社会行为中有趣的话题。在达尔文诞辰一百周年之后,西方科学家们还在不停争论着利他行为的起因和影响。应当注意到的是,文中提到的群体选择的数学公式、出于本能的亲缘选择和对互惠选择的信任都是对动物行为的简单化的模拟,实际的自然中有更多因素需要综合考虑,基因选择模型是最好的理论,成功的解释了利他行为的演化源头。[1]达尔文在The Descent of Man (2nd ed., 1875, pp.187-8)中指出人类的整个历史表明合作精神不但是人的本性,而且比任何自利行为更加根本。如果不是这样,我们将不会看到人类社会的整合和组织在多少世纪和千年以来的增长("the whole of human history show that a co-operative spirit is not only natural to men, but more deeply rooted than any self-seeking tendencies. If this were not so we should not see the growth of integration and organization of his communities which the centuries and the millennia plainly exhibits." )[6]。达尔文的观点很好的印证了中国儒家和道家对人性本善的看法[7][8]。
参考资源
[1]Strong,Eric. The Evolution of Altruism 网址
http://endeavor.med.nyu.edu/~strone01/altruism.html
[2]Stanford Encyclopedia of Philosophy: Biological Altruism 网址
http://www.science.uva.nl/~seop/entries/altruism-biological/
[3]Moor, Jim. The Evolution of Reciprocal Sharing 网址
http://weber.ucsd.edu/~jmoore/publications/Recip.html
[4]www.brooklynparrots.com/labels/Brooklyn%20Parrot.html.
[5]Richard Dawk, The Selfish Gene
[6]The Race Question 网址 http://en.wikipedia.org/wiki/The_Race_Question
[7]《道德经》
[8]《孟子·告子上》