图5. 菲律宾跗猴,曾经被认为属于原猴亚目,现在则普遍认为是简鼻亚目大家族的成员(图片出处:维基百科)
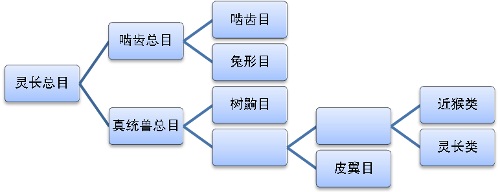
灵长目的分类工作是由瑞典博物学家林奈在1758年首次进行。在他的著作《自然系统》的第十版中,将灵长目分为四个部分:人、猴(包括猿及其他猴子)、狐猴、蝙蝠。在该书1735年的第一版中,他为人、猴等动物用了一个统称:人型动物。1839年,Henri Marie Ducrotay de Blainville模仿林奈的分类方法,建立起了四足目、翼手目等分类方法,但没有为大众所认同。
在早期的分类方法中,灵长类被分为两个科:原猴科和类人猿科。原猴科包括所有的原猴亚目动物,而类人猿科则包括所有的猿类。
1.3 杂交
灵长类通常在被囚禁时进行配种杂交,但偶尔也见其在野外自主进行。融合在两个种族的活动范围交叠时发生,造成这样的区域交叉可能是人为造成的,比如在动物园里所有动物聚集在一起,或者由于诸如人类的掠夺行为造成的环境压力。更高范围上的杂交——属间配种,也曾见于野外。尽管所属的种族已经分离了几百万年之久,在狒狒(Gelada)和北非狒狒(Hamadryas Baboon)之间还是会出现通婚交配。
生理特征
灵长类动物在栖居树木的共同特征下,有许许多多的差异。尽管远古灵长类在进化时有本身独特的身体结构,其后代在漫长的进化过程中不断适应环境,而演变为今日的丰富多彩。它们的各种特征包括保持力需要的胸腔肋骨中的锁骨、保证各个角度自由活动的肩关节、在前端的五指、后肢的宽大脚趾和适合抓握物体的拇指(Opposable thumbs/可对应拇指)、指甲、触觉灵敏的指头(趾头)末端、嗅觉退化的倾向、牙齿数量相对退化、复杂的视觉系统(视觉灵敏和色感)、与躯干不成比例的大脑、扩大的大脑皮层、成对的乳腺、普遍的一胎、较长的妊娠期等。
灵长类动物通常高度社会化,并且有等级之分。新世时期开始灵长类行程一夫一妻制,并形成了稳定的父权社会。
从解剖学角度看
图6. 灵长类动物的头骨大小比较(图片出处:维基百科)
灵长类动物在头部有两只向前的眼睛:这样的双目观察能够提供准确的距离感。眼窝上方有突兀的眉骨,为其他脆弱的脸部骨头提供保护(这些骨头保证了咀嚼时不易疲劳)。Strepsirrhines在眼窝后部有一道活动的横骨,在眼窝内运动并保护眼球。与之对比的,高级的灵长类则进化出了固定在头骨上的眉骨。
灵长类的头骨上有一个巨大的穹顶——头盖骨,这在同类中显得与众不同。头盖骨保护了下面的大脑,同样与众不同。人类的颅腔体积(头骨内的空间大小)比非人类的灵长类动物中最大的颅腔都要大三倍,从中可见人类拥有更大的大脑。人类颅腔体积平均是1201立方厘米,而大猩猩是469立方厘米,黑猩猩是400立方厘米,猩猩是397立方厘米。灵长类进化的首要方向便是大脑,特别是新大脑皮层(大脑皮层的一个部分),它与感官知觉相关:带来了空间感、抽象推理能力、知觉意识、以及——对人类而言——语言文字。在其他哺乳动物严重依靠嗅觉时,栖居树木的灵长类则发展出了高级视觉系统,退化了大脑的嗅觉部分,并增加了复杂的社会行为。
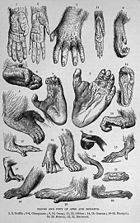
图7. 1893年的一幅画,描绘了灵长类的手和脚(图片出处:维基百科)
灵长类一般在每个前肢各有五个趾头(指头),每个趾的末端都有角质化的趾甲。手足的趾尖皮肤都很敏感,构成了发达的触觉系统。多数灵长类都有适合抓握物体的拇指(opposable thumbs),这是灵长类标志性的特征,但并不独有。例如负鼠便有这样的手指。拇指让动物可以使用工具。对于灵长类,这样的手指配上短小的趾甲是远古时期树丛间行走的产物。由此发展出的许多上肢行走方法(例如用臂在树枝间荡行)则在灵长类的行走方法方面具有重要意义。原猴亚目则在脚上第二趾上有爪型趾甲。
灵长类动物的颈骨在所有圈形胸骨中非常明显,它为肩关节提供了很大的活动空间。猿类的肩关节和手臂有着更多的自由度,则不仅是因为肩胛骨长在背上,更由于拥有与旧世灵长类相比更宽大、扁平的胸廓,以及更短并更少活动的脊椎(有些种类的灵长类由于椎骨底部大幅减少,造成尾骨的消失)。旧世灵长类与猿的不同之处在于它们多数都有尾巴,而与新世灵长类的区别则是它们的尾巴从来也不能抓握。只有新世灵长类拥有可以抓握物体的尾巴。
灵长类的吻部(下颌)显示出缩减的趋势。技术上说,旧世和新世猴类的区分在于鼻部的结构,而旧世猴类和猿的区别在于牙齿的排列。新世猴类的鼻孔面朝侧部,而旧世猴类的鼻孔则朝向下。灵长类动物中有各式各样的牙齿排布方式,并且不论掉了多少门齿,总有至少一颗较低的门齿还保留着。在多数原猴(strepsirhinis)中,较低门齿组成一排被认为食草用的排齿,并且可能有修饰作用。相对于新世猴类的十二颗,旧世猴类同样有八颗前臼齿。旧世灵长类根据臼齿上尖头的数目被分为猿和猴类:猿有五个,而旧世猴类有四个。原始人类的臼齿尖(上次尖)是在灵长类历史早期进化而来的,与此相对应的原始的较低臼齿尖(下前尖)则消失不见。原猴亚目则有其独有的固定上唇、潮湿的鼻尖以及朝内的较低前齿。
灵长类的视力进化与多数哺乳类动物相比都显得与众不同。灵长类的祖先发展出三色视觉(能看到三种颜色),而夜行性动物、恒温动物以及其他哺乳动物的祖先在恐龙时期则丢失了视网膜中三分之一的圆锥细胞。因此只有鱼类、爬行动物和鸟类,以及一些灵长类动物拥有三色视觉,而其他所有的哺乳动物都只能辨认两种颜色和它们的混色。狭鼻类动物(人、猿、猴等)一直保持三色视觉,而阔鼻类动物则不全是,事实上只有很少数量的阔鼻类动物拥有三色视力。
嚎猴(Howler monkeys)则通过将红-绿视蛋白基因自然复制而重新拥有了固定的三色视觉系统。嚎猴是新世猴类中最独特的一种,它们吃树叶为生,偶尔享用些水果。它们偏好消费的树叶(口感嫩、有营养、易消化)则只能够被红绿信号感知。针对嚎猴饮食偏好的田野研究显示,发展出固定的三色视觉是嚎猴接受自然选择的结果。
3.1 性二型性
图8. 性二型性在雄性(灰色)和雌性(棕色)北非狒狒中非常明显(图片出处:维基百科)
同一种类动物中不同性别个体之间的差别称作性二型性。猿类常常显示出这样的特性。总体而言,包括猿类和一些猴类的旧世灵长类比新世灵长类更多的显示出性二型性。近期的研究包括DNA对比检测灵长类二型性的种类变化,以及造成性二型性的基本原因。灵长类的性二型性通常表现在体型、犬齿大小、以及毛皮/皮肤颜色。差异可以归因于不同因素的影响,包括交配制度、种类体型、自然环境以及饮食结构。
比较分析法已经证实了性别选择(雌雄淘汰)假说,并发展出一套更完整的学说来解释灵长类动物性别选择、自然选择和交配制度的关系。研究同时探寻了性二型性进化过程中不同的选择和非选择机制的作用。研究结果表明,性二型性是雌性和雄性的特征共同变化的产物。一些化石证据表明二型性是收敛进化的,并且显示一些已经灭绝的原始人类在性二型性上的差异超过了任何一种灵长类动物。
3.2 运动
图9. 垂直跳跃的王冠狐猴,正准备跳到另一颗树上(图片出处:维基百科)
灵长类动物的运动方式多种多样,有用臂、双足、跳跃、树栖、以及在陆地上用四肢行走、指节爬行等。不少原猴亚目的动物是垂直跳跃并依附在树上的,包括许多灌木猴、所有的大狐猴、以及所有的跗猴(东印度眼镜猴)。原猴亚目的其他种类都是四足攀爬并栖息在树上的,其中一些还能够四足在地面行走,一些则跳跃行走。多数的猴类都同时是爬树和四足陆地行走的高手。长臂猿、蜘蛛猿等则大量运用臂力行走。猩猩使用的是一种类似的运动方式——四足爬行,它们用手臂和下肢在树林间支撑起沉重的身体。而黑猩猩和大猩猩则用指节爬行,并且可以直立进行短距离行走。人类是唯一能够完全直立行走的灵长类动物。
动物行为
4.1 社会系统
图10. 日本短尾猴在温泉中集体沐浴(图片出处:维基百科)
Richard Wrangham 提出,社会系统分类的最好方法是根据集体间雌性活动量的大小。他提出了四个类型:
• 雌性迁移系统-雌性离开所出生的群体。雄性一直留在原来的群体中,而雌性与集体的联系则并不紧密。这样的群体通常很小,常见于黑猩猩社会:雄性黑猩猩会一起行动保卫领地。在新世猴类中,蜘蛛猴和毛蜘蛛猴拥有这样的社会系统。
• 雄性交换系统-雌性保留在原来群体中而雄性则在成年后移居他处。允许多配偶的社会便属于这个类型。这样的社会群体通常会稍大一些,常见于环尾狐猴、卷尾猴等。
• 一夫一妻系统-一个雄性和一个雌性的稳定结构,有时还伴有后嗣。照顾家庭和服务社会(如保卫领土)的工作都是双方共分的。子女成年后会离开父母的领地。这样的社会比较典型的是长臂猿群体。不过,一夫一妻制在这里并不代表性生活的忠贞。
• 孤独型(对雌性来说)-雄性会一起保卫自己的领土,而这其中会包括几个雌性的活动地区。此种结构被发现于猿类。猩猩不怎么关心领土事务,但其群体也是这样的组织类型。
其他类型的社会系统也有。例如,嚎猴的雄性和雌性都可能在成年后从原有群体中转移到别处,造成的结果便是群体内的成员,无论雌雄,相互都没有亲密的血缘关系。
Attach:Primates_picture11.jpg Δ
图11. 黑猩猩是集体动物(图片出处:维基百科)
灵长类的专家珍•古道尔博士曾在坦桑尼亚的固北河国家公园内研究,发现了黑猩猩社会的分-合特征。白天整个群体为了进食而分散开来,夜间则聚集在一起休息。这样的社会结构同样见于北非狒狒、蜘蛛猴和倭黑猩猩。狒狒的社会系统也与之类似:许多较小的群体会聚在一起组成最高600名成员的临时集体。
这些社会系统主要由三个因素影响:资源、群体大小、掠夺行为。一个社会群体之内既有合作也有竞争。合作会以相互梳理毛发的形式体现:互相清除皮肤上的寄生虫、清洁伤口、分享食物,也会出现集体抵御侵略和掠夺的活动。而竞争则在食物、居住地点和配偶的问题上显现出来。竞争的结果便是建立起了社会等级。
4.2 种间联系
不同种类的灵长类会相互合作。主要内容便是共同抵御外部掠夺。抵御时还会出现分工合作,例如负责发警报。典型的便如赤尾猴和包括疣猴、白腹长尾猴等的其他种族合作。它们的共同敌人通常是黑猩猩。
又如在南美洲,松树猴会与卷尾猴联合起来。不过这种联合更多的是为了食物,而非抵御侵略。
4.3 认知和交流
狐猴、懒猴、跗猴和新世猴类都依赖嗅觉信号进行各种社会行为和繁殖活动。它们的鼻子能够分辨出特殊的标记气味,这些气味是被腺体分泌出来标明领地用的。这个行为便是这些灵长类动物相互交流的主要方式。旧世猴类和猿类中,嗅觉能力则普遍退化,而视觉则成为主要信息感知器官。灵长类动物还利用声音、动作、面部表情来展现心理状态。
4.4 生命阶段
灵长类比其他哺乳动物的生长更慢。除了人类,所有的灵长类幼崽都依靠母乳喂养传递养分,并由母亲保护、提携。某些种类也有雄性,特别是父亲,负责子女的活动和安全。其他家庭成员,包括兄弟姐妹或是妯娌,也会参与照看幼崽。多数的灵长类雌性在哺乳期会停止排卵,而在停止哺乳后会重新拥有生殖能力。这常常会造成母子之间的冲突:幼崽往往不愿意断乳,希望继续食用。
灵长类比同等体型的哺乳动物拥有更长的幼年(从断乳到性成熟)。未成年的灵长类相对于成年的更容易因为竞争而遭受饥饿,因此它们在这个阶段里学习如何防止食物被掠夺。它们在这个阶段还会学习社交和打斗,这些知识通常都是在玩耍中获得的。
成熟期较晚给灵长类带来了与同等体型哺乳动物相比更长的寿命。并且雌性的平均寿命要长于雄性。
4.5 饮食
图12. 正在吃树叶的非洲黑白疣猴(图片出处:维基百科)
灵长类的食物来源非常广泛。多数灵长类都吃水果,以此吸收易消化的碳水化合物和油脂作为能量。然而灵长类也需要其他食物,例如昆虫或者树叶,以获取氨基酸、维生素和矿物质(微量元素)。许多灵长类都有特殊的、支持特定食谱的生理结构。例如以树叶为食的嚎猴、黑白疣猴和狐猴等,消化道都稍长一些,以便从不易消化的树叶中吸取更多营养。又比如黑颊白眉猴的牙齿上有一层厚厚的牙釉质,帮助它们对付硬壳的水果和坚果。
狒狒是唯一一种主要食草的灵长类。跗猴是食肉最多的灵长类——专吃昆虫、爬行动物、和一些其它动物。普通黑猩猩则吃几乎所有能吃的东西,包括从别的灵长类(例如红疣猴)那儿抢来的食物。
栖息地及分布状态
图13. 在印度北部城市阿格拉的恒河短尾猴(图片出处:维基百科)
灵长类是从树栖动物进化而来,而灵长类本身的确有很多都是一生在树上度过的。但另一些种类,例如狒狒等,则是部分甚至全部时间都在陆地上生活的,例如人类。除去人类不算,灵长类生活在非洲、印度、东南亚和南美洲的热带/亚热带雨林、树林等许多林区中。也有一些灵长类动物生活在非热带地区,比如在本州岛北部生活的日本短尾猿,栖居于一年有八个月覆盖着白雪的山上。而它的近亲-北非短尾猿则生活在阿尔及利亚和摩洛哥的阿特拉斯山中。不同灵长类动物居住的纬度也有很大差异:黑扁鼻猴被发现生活在海拔4700米的横断山脉,在海拔5000米的埃塞俄比亚高地也出现过狒狒的身影。尽管多数灵长类水性都不佳,仍然有一些种类,包括长鼻猴等,是生活在湿地和近水区域的游泳高手。它们的趾间已经进化出了小小的边,起着蹼的作用。有的灵长类,诸如恒河短尾猿和灰叶猴,可以适应经过人类改造的环境,甚至生活在城市里。
与人类的关系
灵长类动物和人类有许多不同的关系。有些被收养为宠物,有些被作为实验室的生物样本,有的甚至替代宇航员被送上太空。有的灵长类还会被作为服务性动物,为残障人士提供帮助。
灵长类会传播许多传染疾病,例如疱疹、麻疹、埃博拉病毒、狂犬病、马尔堡病毒以及肝炎病毒。其中的一些能够被传染给人类,包括潜在致命的疱疹B型病毒。但话说回来,人类行为从来都是对灵长类不利的,并且已经造成了许多灵长类物种濒临灭绝。
6.1 在科学实验中的角色
图14. 被关在板条箱中进口来,将用在实验中的猴子(图片出处:维基百科)
全世界有数千种非人类灵长类动物(NHPs)被用来做动物科学的研究。由于与人类的基因相似性,灵长类是做与人类相关的研究时绝佳的生物样本。短尾猿(尤其是恒河短尾猿)、非洲绿猴、黑猩猩、狒狒和长尾猴,无论是家养的还是野生的,都被大规模用于动物实验。通常灵长类动物实验的内容包括潜伏期测试、神经实验、眼科研究等。对NHP的解剖,特别注重在其与人类极其相似的部位:大脑和眼睛。2004年欧盟使用了大约10000只动物;2005年仅仅英国就对3115只非人类灵长类动物做了4652次动物实验。2004年,共有3100只非人类灵长类动物被囚禁在美国的动物园、马戏团和实验室里。其中1280只曾被用来做动物实验。许多国家的政府都对灵长类实验有特殊的严格规定。在美国,联邦政府对非人类灵长类动物的居住、饮食、数量和驯养等我各方面都做出了相应规定。欧洲的一些活动团体,例如欧洲结束动物实验联合会,正在寻求发布一项禁止一切灵长类动物实验的禁令,以回应欧洲现有的关于动物实验的法律法规。
6.2 法律地位
在整个灵长类下,只有人类被认为是拥有人性的个体,并被联合国人权宣言和其它法律所保护。非人类灵长类动物的地位富有争议。诸如“大猩猩计划”的项目和组织,以及诸如Ignaas Spruit(保护灵长类组织的总干事)的人,都在极力推动增加非人类灵长类动物的法律地位。
6.3 物种保护
图15. 濒临灭绝的苏门答腊猩猩(图片出处:维基百科)
国际自然保护协会(IUCN)指出,有至少三分之一的灵长类动物的生存正在受到严重威胁。一般来说这样的威胁包括森林退化、消失,食物源减少,以及人类为了医学或食用而对灵长类的捕杀行为。森林的锐减被认为是威胁灵长类动物比较主要的原因之一,因为超过90%的灵长类都生活于此。森林的消失是由于农业开垦造成的,其它导致森林减少的因素还包括工业伐木、生活用木料的采集、开矿以及筑坝。在印度尼西亚,为了增加棕榈油产量,大片的低海拔森林被砍伐殆尽。1998到1999年间的卫星图像分析表明,平均每年有1000只苏门答腊猩猩从吕宋岛的生态系统中消失。
由于体型较大(大于5千克),需要更多食物的灵长类动物便比其它小动物有更多潜在的生存危机。又因为灵长类的寿命长,性成熟晚,代与代间隔的时间相对比较长,这造成了种群数量一旦遭到诸如捕猎一类活动的严重打击,很难在短时期内恢复到原有水平。一些非洲城市则被估计有将近一半的蛋白质消费是从灵长类得来的。对一些濒危动物的捕猎,诸如长尾猴等,早已超过了可以承受的界限。屡禁不止的原因是,这些动物体型大,易于运输的同时能够保证平均的高利润。另外由于农业的发展侵蚀了灵长类的森林植被,灵长类便以农作物为食,破坏农民的收入。灵长类这样的行为给当地人留下负面印象,长远地阻碍了灵长类保护工作的发展。
拥有五种独有灵长类的马达加斯加,在最近几年里已经见证了物种大面积的消失。自从人类在1500年前定居这里,至少8个类,15种灵长类动物都由于人类猎杀和栖息地消失而灭绝。
在亚洲,印度教、佛教和伊斯兰教都禁止食用灵长类,不过仍然有灵长类被猎杀捕食。一些范围小的传统信仰允许这样的行为。饲养宠物和传统医学也在另一个侧面增加了民间捕猎灵长类动物的需求。最近几年,中国日益增长的中产阶级也开始钟爱于富有异国情调的小宠物。但也有例外:曾经被作为动物实验模板的恒河短尾猿,在经过1960年代过量捕猎威胁到群体数量时,被人做出规定保护起来。这项政策极其有效,以至于如今大量的恒河猴被视作有害物。
在中南美洲,森林减少和捕猎是当地灵长类的两个主要威胁。中美洲现在已经鲜见有大片的树林了。这让森林更易受到比如农场的影响,并且会造成空气湿度降低和植被的变化。可生活区域的限制造成灵长类动物大量近亲繁殖,更进一步形成了种群发展的瓶颈。不然为何会有一半数量的灵长类消失呢?
图16. 濒临灭绝的大猩猩(图片出处:维基百科)
对美国而言,主要的问题是宠物贸易。尽管1975年政府已经禁止了灵长类的进口,在美国-墨西哥边境上还时有发生走私的事情。从3000美金的猴子到30000美金的猿,价格不尽相同。
美国濒临灭绝的灵长类共有21种,其中8种同时也在IUCN所列“25种世界濒危灵长类”中。其中一种红疣猴在1978年最后一次被人看见,1993-99年无人发现其任何踪迹,直到2000年正式宣布绝种,也成为了21世纪第一个灭绝的灵长类。
参考资源
(1)维基百科"Primates"条目:http://en.wikipedia.org/wiki/Primates
(2)百度百科"灵长目"条目:http://baike.baidu.com/view/33310.htm
(3)北京动物园猩猩馆
(4)
(5)
(6)http://www.cnki.com.cn/Article/CJFD1994-SWXT405.022.htm














{kind=link}